Research article
Changes in apoptotic factors in hypothalamus and hippocampus after acute and subchronic stress induction during conditioned place preference paradigm
Abbas Haghparast1[*],2,3, Zahra Fatahi1, Shabnam Zeighamy Alamdary1, Fariba Khodagholi1
1Neuroscience Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran2Iran National Science Foundation, Tehran, Iran
3Neurobiology Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran
EXCLI J 2013;12:Doc1001
Abstract
The hypothalamus (HYP) and hippocampus (HIP) are important regions involved in stress responses. These areas are also associated with reward processing. In this study, the effects of acute and subchronic stress on the changes in apoptotic factors (Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation) in the HYP and HIP during conditioned place preference (CPP) paradigm were evaluated. Male Wistar rats were divided into two saline- and morphine-treated supergroups. Each supergroup contained control, acute stress (AS) and subchronic stress (SS) groups. In all groups, CPP paradigm was done and thereinafter alterations of apoptotic factors were measured by western blot. The results revealed that in the HYP, all mentioned factors increased significantly in saline- or morphine-treated animals during AS and SS. On the other hand, in the HIP, Bax/Bcl-2 ratio in saline-treated animals increased significantly during AS and SS, while in morphine-treated animals this ratio did not have any significant alteration during AS and was decreased during SS compared with morphine-control group. Caspase-3 and PARP increased during AS and SS in saline- or morphine-treated animals. For example, caspase-3 increased during AS and SS in morphine-treated animals by 2.4 folds and PARP (89 KDa) increased by 3.1 and 3.5 folds, respectively. Interestingly, the increase of apoptotic factors in morphine-treated animals was more considerable than that of saline-treated animals. These results strongly implied that AS and SS trigger apoptotic events in the HYP and HIP of saline- and/or morphine-treated animals as well as morphine reinforces the effect of stress on the induction of apoptosis.
Keywords: apoptosis, hypothalamus, hippocampus, forced swim stress, conditioned place preference, rat
Introduction
Stressful events cause a strong set of behavioral, hormonal, cellular and molecular responses that assist organisms in adapting to the physical and social environment (Trainor, 2011[42]). The hypothalamus (HYP) and hippocampus (HIP) have been shown to be the important regions involved in response to stressor (Lucas et al., 2011[30]). The HYP is the center of stress response, also an area of the brain that integrates various stress pathways (Lee et al., 2005[25]). In response to stress, the HYP is activated and triggers hypothalamic-pituitary-adrenal (HPA) axis activation; the paraventricular nucleus of the HYP release corticotrophin releasing hormone into the hypothalamic portal system at the median eminence that results in increased release of adrenal corticotrophic hormone (ACTH) into the systemic circulation. ACTH is transported to the adrenal gland where it causes the adrenal cortex to increase the synthesis and release of glucocorticoids hormones including corticosterone in rodents and cortisol in primates (Lovallo, 2006[29]; Zhu et al., 2001[45]). Moreover, the HYP seems to play an important role in the physical signs of drug-induced withdrawal (Prieto-Gomez et al., 1984[35]). Lateral hypothalamus is involved in reward processing and addictive behaviors (Aston-Jones et al., 2010[3]). The HIP which has a principle role in learning, memory and emotional processing (Cui et al., 2010[11]; Fuchs and Flugge, 2003[16]), has been shown to be involved in the regulation of the HPA axis activation and the behavioral responses to stress (Cai et al., 2008[7]). In addition, the HIP is also associated with morphine dependence and previous studies demonstrated that this area of the brain is involved in morphine-induced conditioned place preference and withdrawal behavior (Yang and Pu, 2009[43]).
Apoptosis is a form of cell death that occurs through the activation of intracellular suicide mechanisms in response to deleterious stimuli such as stress, addictive drugs, growth factor withdrawal, etc (Sreedhar and Csermely, 2004[40]; Zhang et al., 2004[44]). This process is regulated by antiapoptotic (Bcl-2) and pro-apoptotic (Bax) effector molecules. Briefly, Bax is a soluble protein present predominantly in the cytosol whereas Bcl-2 is present in mitochondria and functions as a repressor of apoptosis. During the induction of apoptosis, Bax shifts to mitochondrial membranes and causing changes in mitochondrial permeability that leads to release of apoptotic protein cytochrome C into the cytoplasm (Li et al., 2010[26]). Subsequently, release of cytochrome C results in an apoptosome transactivation of the complex procaspase-3 to active caspase-3 (one of the most important factors for occurrence of cells death) (Haack et al., 2008[19]; Zlatkovic and Filipovic, 2012[46]). It has been clarified that opiates such as morphine and heroin are capable to induce apoptosis in different kinds of the neural cells including neuron, microglia and astrocyte (Kim et al., 2001[22]; Hu et al., 2002[21]). Exposure to morphine strongly induces apoptosis in the parietal, frontal, temporal and occipital regions of the rat brain (Chen et al., 2008[9]; Fan et al., 2003[15]). Moreover, it was shown that stress leads to atrophy and functional impairments in several key brain areas including the cerebral cortex, amygdala and HIP. Evidence also indicates that prolonged stress inhibits growth and survival mechanisms at the cellular level and leads to apoptosis (Haack et al., 2008[19]). Therefore, in the present study, we aimed to investigate the effects of acute and subchronic stress in saline- or morphine-treated rats in conditioned place preference paradigm on the alteration of apoptotic factors (Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation) in the hypothalamus and hippocampus areas.
Materials and Methods
Animals
Adult male Wistar rats (Pasture Institute; Tehran, Iran) weighing 230-280 g were used in this study. The animals were kept in an animal house with a 12-h light/12-h dark cycle with controlled temperature (22 ± 2 °C) and ad-lib access to food and water. All animals were allowed to adapt to the laboratory conditions for at least 1 week before experiment and were handled for 5 min/day during this adaptation period. All experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health Publication No. 80-23, revised 1996) and were approved by the local ethical committee, Shahid Beheshti University of Medical Sciences.
Materials
The materials used in this study were morphine sulfate (Temed Co., Tehran, Iran), antibodies directed against Bax (Cat No. #2772), Bcl-2 (Cat No. #2876), caspase-3 (Cat No. #9665), PARP (Cat No. #9542), β-Actin (Cat No. #4967), secondary antibody IgG (Cat No. #7074), Cell Signaling Technology, USA) and Electrochemiluminescence (ECL) kit (Amersham Bioscience, USA). Morphine was dissolved in 0.9 % sterile saline at a concentration of 5 mg/ml and injected subcutaneously (s.c.).
Conditioning apparatus and paradigm
The rewarding properties of morphine were investigated using a conditioned place preference (CPP) paradigm. Place conditioning boxes were made of Plexiglas and consist of two sides (30 cm × 30 cm × 40 cm), but differed in shading and texture. Compartment A was white with black horizontal stripes 2 cm wide on walls and also had a net-like floor. Compartment B was black with vertical white stripes, 2 cm wide and also had a smooth floor. The third compartment, C, was a red tunnel (30 cm × 15 cm × 40 cm). It protruded from the rear of the two large compartments and connected the entrances of them. In this apparatus, rats showed no consistent preference for either large compartments (A and B), which supports our un-biased CPP paradigm. It took place in the following manner.
Pre-conditioning phase
On the first day, each rat was placed separately into the apparatus for 10 min with free access to all compartments (A, B and C), and the amount of time spent in each compartment was measured to assess unconditioned preference. (The position of the animal was defined by the position of its front paws.) In the particular experimental setup used in the present study the animals did not show a significant preference for either of the compartments (A and B) during preconditioning phase.
Conditioning phase
Place conditioning phase started 1 day after preconditioning phase for 3 days. This phase consisted of six 30-min sessions (three saline-treated and three morphine-treated). These sessions were conducted twice each day (days 2-4) with a 6-h interval. On each of these days, separate groups of animals received one conditioning session with morphine and one with saline. During these sessions, the animals were confined to one compartment by closing the removable doors. Animals of each group were injected with morphine or saline and immediately confined to one compartment of the apparatus. After 6-h interval, following administration of saline, the animals were confined to the other compartment. Treatment compartment and order of presentation of morphine and saline were counterbalanced for either group.
Post-conditioning phase
On the fifth day, the wall was removed, and each animal was allowed free access to all apparatus. The mean time spent for each rat in both large compartments was recorded by ethovision software (Version 3.1), a video tracking system for automation of behavioral experiments (Noldus Information Technology, the Netherlands). Conditioning score (CPP score) represents the time spent in the reward-paired compartment minus the time spent in the same compartment on the pre-conditioning phase during a 10-min period.
Forced swim stress
The forced swim stress (FSS) was performed in a Plexiglas tank (50 cm height × 30 cm diameter) filled with 35 cm depth of water (24 ± 27 °C). Each rat was forced to swim individually for 6 min once a day. For induction of acute stress (AS), the animals received FSS for 6 min just one day but, in subchronic stress (SS) groups, the animals received FSS for 6 min once a day for 3 consecutive days.
Western blot analysis
For Western blot analysis, three rats per group were decapitated after performance of the CPP paradigm and the brains were rapidly removed on ice. The brain was placed in a stainless steel brain matrix, and the hypothalamus and hippocampus were dissected bilaterally according to the atlas of Paxinos and Watson (2007[34]). All tissues were placed into liquid nitrogen to be frozen. Tissues were sonicated in 1 % sodium dodecyl sulfate buffer in Tris-EDTA, pH 7.4, containing 1× protease inhibitor cocktail, 5 mM NaF, and 1 × phosphatase inhibitor cocktail. Samples were boiled for 5 min and centrifuged at 16,100 × g for 10 min. The proteins were electrophoresed in 12% SDS-PAGE gels, transferred to poly vinylidene fluoride membranes. Blots were incubated in blocking buffer (10 % nonfat dry milk powder in Tris-buffered saline containing 0.5 % Tween-20, TTBS) for 1 h at room temperature (RT), were incubated with primary antibody (1:1000 dilution) overnight at 4 °C and then washed 10 min × 3 in TTBS. Blots were incubated with secondary antibody IgG (1:3000 dilution) for 1 h at RT, washed 10 min × 3 in TTBS, treated with ECL reagents and exposed to film. Data analysis was performed by Image J., measuring integrated density of bands after background subtraction. Protein concentrations were determined according to Bradford's method (Bradford, 1976[5]).
Experimental design
Male Wistar rats were divided into two main supergroups, saline- and morphine-treated animals. Each supergroup consists of control, AS and SS groups. In all of the groups (n = 6-8/group for behavioral studies and 3-4/group for molecular studies), the animals passed CPP stages according CPP protocol. In saline-treated animals, control group received saline (1 ml/kg) during the conditioning phase without any stress, AS group received saline during the conditioning phase and also FSS on the post-conditioning phase10 min before was put on apparatus and SS group received FSS during 3-day schedule conditioning phase, 10 min before saline injection once a day. In morphine-treated animals, control group treated with morphine (5 mg/kg) during the conditioning phase, AS group received morphine in the conditioning phase, also FSS on the post conditioning phase 10 min before was put on apparatus and SS group received FSS during the conditioning phase 10 min before administration of morphine. Conditioning score was calculated for each rat on the test day.
For measuring the alterations of apoptotic factors, on the fifth day, the animals were killed and the brains were removed. In each animal the HYP and HIP were immediately dissected out and delivered into liquid nitrogen. Tissues were prepared for Western blot analysis. In each area, the level of Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation were evaluated.
Statistics
All data were shown as mean ± SEM (standard error of mean) and in order to compare the conditioning scores in control and experimental groups, one-way analysis of variance (ANOVA) followed by post hoc Newman-Keuls multiple comparison test was used. In molecular section the optical densitometric data were analysis by two-way analysis of variance (ANOVA) followed by post hoc Bonferroni test or Newman-Keuls multiple comparison test are needed, respectively. The level of statistical significance was set at P<0.05 and calculations were performed by using GraphPad Prism (Version 5.0) software.
Results
Effects of acute and subchronic stress on the change in conditioning scores in CPP paradigm
For determining the effects of acute and subchronic stress on the change in conditioning scores, the animals passed CPP stages according to CPP protocol and afterward, conditioning score was calculated. One-way ANOVA followed by Newman-Keuls test [F(5,42)=4.336, P=0.0033] clarified that in saline-treated animals, conditioning scores did not have any alteration in respective saline-control group after application of acute stress but it decreased after performance subchronic stress. On the other hand, in morphine-treated animals, conditioning scores decreased after application of both acute and subchronic stress (Figure 1(Fig. 1)).
Change in apoptotic factors in the hypothalamus after induction of acute and subchronic stress during conditioned place preference paradigm
In the first step of experiments, to evaluate the effects of acute and subchronic stress on the alternations of apoptotic factors in the HYP in saline- or morphine-treated animals, the CPP paradigm was done and afterward the alterations of Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation were measured by western blot analysis (Tables 1(Tab. 1) and 2(Tab. 2)). Two-way ANOVA followed by Bonferroni test for Bax/Bcl-2 ratio [Morphine treatment effect: F(1,12) = 61.75, P<0.0001; Stress effect: F(2,12) = 35.04, P<0.0001; morphine treatment × stress interaction: F(2,12) = 0.61, P = 0.2857; Figure 2(Fig. 2)], caspase-3 activation [Morphine treatment effect: F(1,12) = 58.27, P<0.0001; Stress effect: F(2,12) = 36.53, P<0.0001; morphine treatment × stress interaction: F(2,12) = 4.44, P<0.0001; Figure 3(Fig. 3)] and for PARP degradation [Morphine treatment effect: F(1,12) = 15.33, P = 0.0031; Stress effect: F(2,12) = 60.49, P<0.0001; morphine treatment × stress interaction: F(2,12) = 10.61, P = 0.0313; Figure 4(Fig. 4)] showed that all measured apoptotic factors increased significantly in saline- or morphine-treated animals during both AS and SS conditions.
Change in apoptotic factors in the hippocampus after induction of acute and subchronic stress during conditioned place preference paradigm
In another set of experiments, we examined the effects of acute and subchronic stress on the alterations of apoptotic factors including Bax/Bcl-2 ratio, caspase-3 and PARP in the HIP by using western blot analysis (Tables 3(Tab. 3) and 4(Tab. 4)) after performance of the CPP paradigm. As shown in Figure 5(Fig. 5), Bax/Bcl-2 ratio in saline-treated animals increased significantly during AS and SS, while in morphine-treated animals this ratio did not have any significant alteration compared with morphine-control group during AS and decreased during SS despite its increase in respective saline-control group [Morphine treatment effect: F(1,12) = 88.06, P<0.0001; Stress effect: F(2,12) = 4.24, P<0.0001; morphine treatment × stress interaction: F(2,12) = 7.59, P<0.0001]. Two-way ANOVA followed by Bonferroni test for caspase-3 activation [Morphine treatment effect: F(1,12) = 79.1, P<0.0001; Stress effect: F(2,12) = 13.51, P = 0.0007; morphine treatment × stress interaction: F(2,12) = 1.59, P= 0.2346] showed that activated caspase-3 increased during AS and SS in both saline- or morphine-treated animals however its increase in morphine-treated animals during AS was not significant (Figure 6(Fig. 6)). Furthermore, PARP degradation increased significantly in saline- or morphine-treated animals during AS and SS [Morphine treatment effect: F(1,12) = 67.38, P<0.0001; Stress effect: F(2,12) = 29.38, P<0.0001; morphine treatment × stress interaction: F(2,12) = 1.98, P = 0.0034; Figure 7(Fig. 7)].
Discussion
The most relevant result of this study is that acute and subchronic physical stress increased the level of apoptotic factors (Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation) in the hypothalamus and hippocampus in saline- or morphine-treated animals and this increase in morphine-treated animals was more noticeable than that of saline-treated animals. In this study, for the induction of stress we used forced swim stress because it is inescapable physical stress and previous studies have shown that this model of stress leads to release of ACTH and corticosterone. Both hormones increased significantly up to 40 min after stress (Abel, 1994[1]; Rittenhouse et al., 2002[36]; Shishkina et al., 2010[38]). Our results showed that AS and SS increased Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation significantly in the HYP in saline- or morphine-treated animals. On the other hand, in the HIP, in saline-treated animals all mentioned factors enhanced significantly during AS and SS. However, in morphine-treated animals Bax/Bcl-2 ratio did not have any significant alteration during AS and decreased during SS compared with morphine-control groups, caspase-3 activation enhanced during AS and SS but its increase during AS was not significant and PARP degradation increased significantly in both AS and SS groups. Previous studies suggested that apoptosis is promoted by overexpression of Bax versus Bcl-2 (Ding et al., 2010[13]). Bax oligomerizes in the mitochondrial membrane and causing the permeabilization of the mitochondrial membrane followed by the leakage of cytochrome C to cytosol (Li et al., 2010[26]; Sreedhar and Csermely, 2004[40]). Cytochrome C activates caspase-3 which, in turn, induces the cleavage of vital cellular substrates including the nuclear enzyme PARP (an enzyme involved in DNA repair). PARP (abundantly present in neurons and glia cells) is cleaved into specific 89-kDa and 24-kDa fragments, which greatly deactivate PARP function as a DNA-repairing enzyme and are as markers of cells undergoing apoptosis (Alvaro-Bartolome et al., 2011[2]; Goebel, 2009[18]). In the current study, the results implied that in saline-treated animals, AS increased the level of all cited factors in the HYP and HIP noticeably. In agreement with our results, Heine et al. (2004[20]) reported that AS increased apoptosis and suppressed new cell birth in the rat HIP; conversely, Haack et al. (2008[19]) revealed that acute restrained stress could not enhance the cortical levels of Bax. Also, there is a study indicating that acute cold stress at 4 °C or acute immobilization stress did not increase apoptotic factors in the HIP and prefrontal cortex (Zlatkovic and Filipovic, 2012[46]). It seems that these different results are due to stress protocol. However, it needs more investigations.
In addition, in these animals, SS increased all stated factors in the HYP and HIP, too. These findings are consistent with other studies showing that chronic stress can suppress growth and survival mechanisms at the cellular level, resulting in neuronal atrophy and apoptosis in different regions of the brain including the HIP, amygdala and prefrontal cortex (Ding et al., 2010[13]; Gianaros et al., 2007[17]; Haack et al., 2008[19]). Evidence also indicates that chronic isolation stress increases Bax/Bcl-2 ratio in prefrontal cortex and leads to the detrimental release of cytochrome C from the mitochondria (Djordjevic et al., 2012[14]; Terashvili et al., 2008[41]). Consistently, Bachis et al. (2008[4]) revealed that chronic unpredictable mild stress increased caspase-3 activation in the cerebral cortex neurons and also Zlatkovic and Filipovic (2012[46]) reported that Bax/Bcl-2 ratio in the prefrontal cortex increased significantly followed by chronic isolation stress. Moreover, behavioral results in these animals explained that, conditioning score decreased during subchronic but not acute stress. It was reported that stress produces conditioned place aversion (Land et al., 2009[24]) and chronic exposure to mild unpredictable stress reduces or abolishes the acquisition of place preference conditioning (Papp et al., 1992[33]). Then, it was deduced that repeated (but not single) exposure to swim stress could induce place aversion.
In other part of our experiments, we showed that in morphine-treated animals, AS and SS evinced apoptotic events in the HYP and HIP in respective control group. Apoptotic factors enhanced significantly in morphine-control group compared with saline-control group, interestingly. These findings eminently confirmed the role of morphine on the induction of apoptosis; nevertheless there are also some evidence emphasizing neuroprotective role of morphine. In agreement with our results, previous studies demonstrated that addictive drugs, such as morphine, can induce apoptosis in cultured neuronal cell lines (Lim et al., 2005[27]) as well as other cells including immune cells (Hu et al., 2002[21]). Hu et al. (2002[21]) revealed that proapoptotic regulatory proteins such as Bax increase in morphine-treated cells. It was also shown that heroin and cocaine decrease mitochondrial potential and elicit an increase in cytochrome C release and caspase-3 activation leading to apoptosis in rat cortical neurons (Cunha-Oliveira et al., 2010[12]). In contrast, Chi et al. (2010[10]) reported that opioid preconditioning induces opioid-receptor-dependent neuroprotection against cerebral ischemia. Also, Kim et al. (2001[22]) showed that morphine protects primary rat astrocytes from apoptosis mediated by sodium nitroprusside. Then, it can deduce that apoptotic or neuroprotective effects of morphine are dose-dependent and used dose in this study (5 mg/kg) has apoptotic effect. In this supergroup, AS increased the level of apoptotic factors in both areas. The enhancement of Bax/Bcl-2 ratio and caspase-3 in the HIP were not significant while in this area PARP increased noticeably then it could propose that AS increased apoptosis in these regions compared with morphine-control group. On the other hand, during SS apoptotic proteins were enhanced in the HYP notably. In HIP, Bax/Bcl-2 ratio was decreased as compared with morphine-control group but caspase-3 activation and PARP degradation increased remarkably. Therefore, it could suggest two possibilities. First, in addition to Bax, some other apoptotic factors such as Bak (Matthews et al., 2012[32]) was responsible for caspase-3 activation in the HIP and second, Bax/Bcl-2 ratio (in both AS and SS group) and caspase-3 (in AS group) increased significantly during the induction of stress but they were returning to the basal level when we dissected out the tissues. Interestingly, the enhancement of apoptotic factors during AS and SS in morphine-treated animals was more remarkable than that of saline-treated animals. It is well known that in addition to stress, morphine activates the HPA axis, leading to an increased release of adrenal corticosterone (Kim et al., 1999[23]). Corticosterone stimulates apoptosis in different kinds of the cells such as primary cultured rat hippocampal neurons, bone cells and also different areas of the brain such as the HIP, amygdala and nucleus accumbens (Liu et al., 2011[28]; Silvestrini et al., 2000[39]). More, it has been shown that glucocorticoid and opiate receptors exist most notably in cell bodies within the HYP and HIP (Cui et al., 2010[11]; Mantsch and Goeders, 2000[31]; Prieto-Gomez et al., 1984[35]) and the activations of these receptors stimulate apoptosis (Cerqueira et al., 2008[8]). Taken together, these findings raised the possibility that in morphine-treated animals, morphine reinforces effect of stress on the increase of apoptotic factors. In these animals induction of both acute and subchronic stress decreased conditioning scores. Provided evidence showed that stress leads to release of dynorphin, this neuropeptide binds to kappa opioid receptors and whereby activates ERK/CREB pathway (Bruchas et al., 2010[6]). ERK signaling in the nervous system has a critical role in memory formation and adaptive responses to changing environments (Shiflett and Balleine, 2011[37]). Therefore, it could suppose that ERK/CREB pathway resulted in memory formation and consequently aversion to the compartment which the animal placed there after receiving stress. However, it needs more studies.
In conclusion, this study showed that AS and SS stimulate the increase of apoptotic factors (Bax/Bcl-2 ratio, caspase-3 activation and PARP degradation) in the HYP and HIP of saline- and/or morphine-treated animals and because of correlative effect of stress and morphine on the induction of apoptosis, this increase in morphine-treated animals is more considerable.
Acknowledgement
This work was supported by the grant (No. 91003540) from Iran National Science Foundation, Tehran, Iran.
References
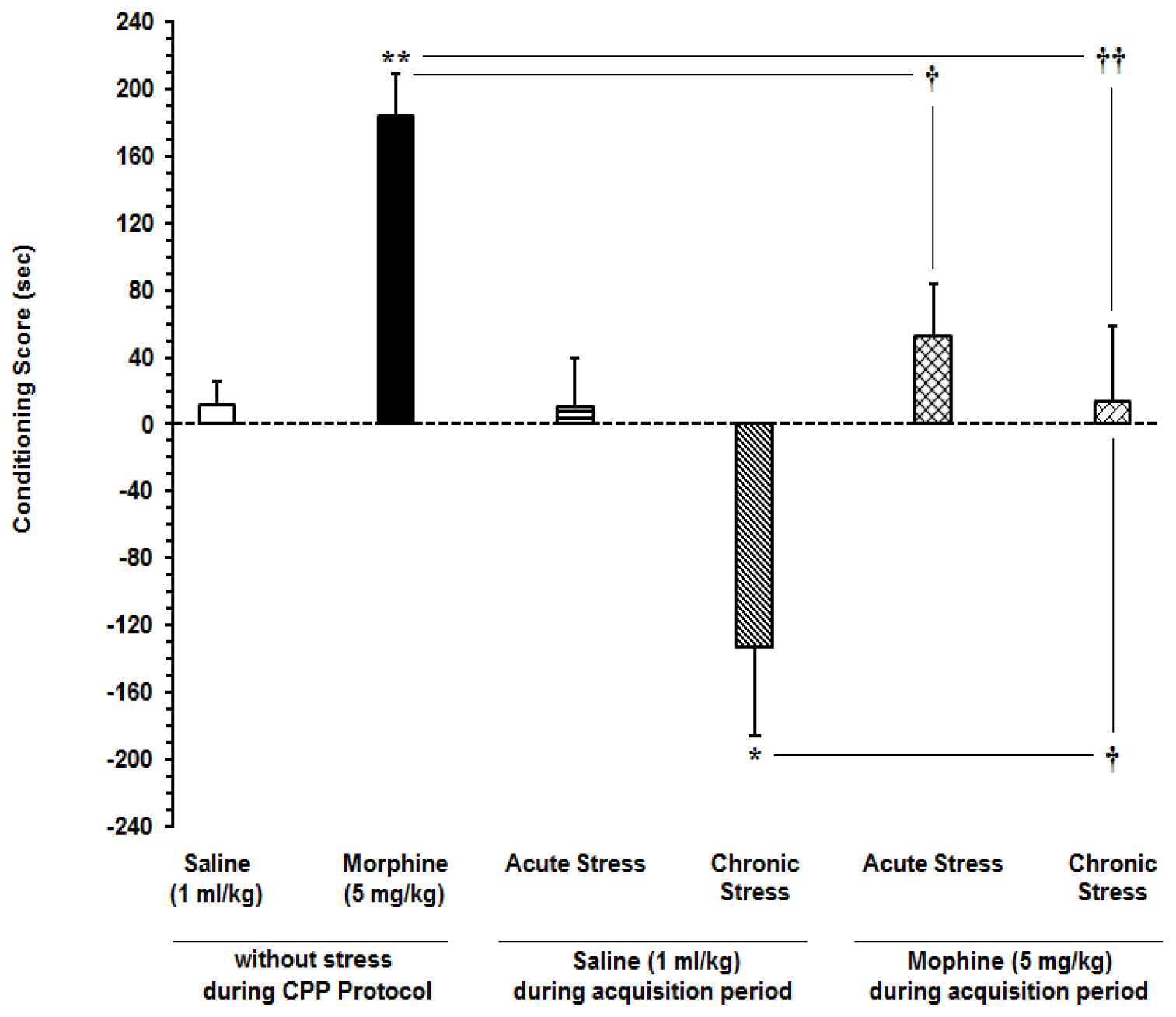
Figure 1: Effects of acute and subchronic stress on the change in conditioning score in conditioned place preference model. Newman-Keuls test; mean ± SEM; n = 6-8 per group
* P<0.05, ** P<0.01 different from the saline-control group
† P<0.05, †† P<0.01 different from the morphine-control group
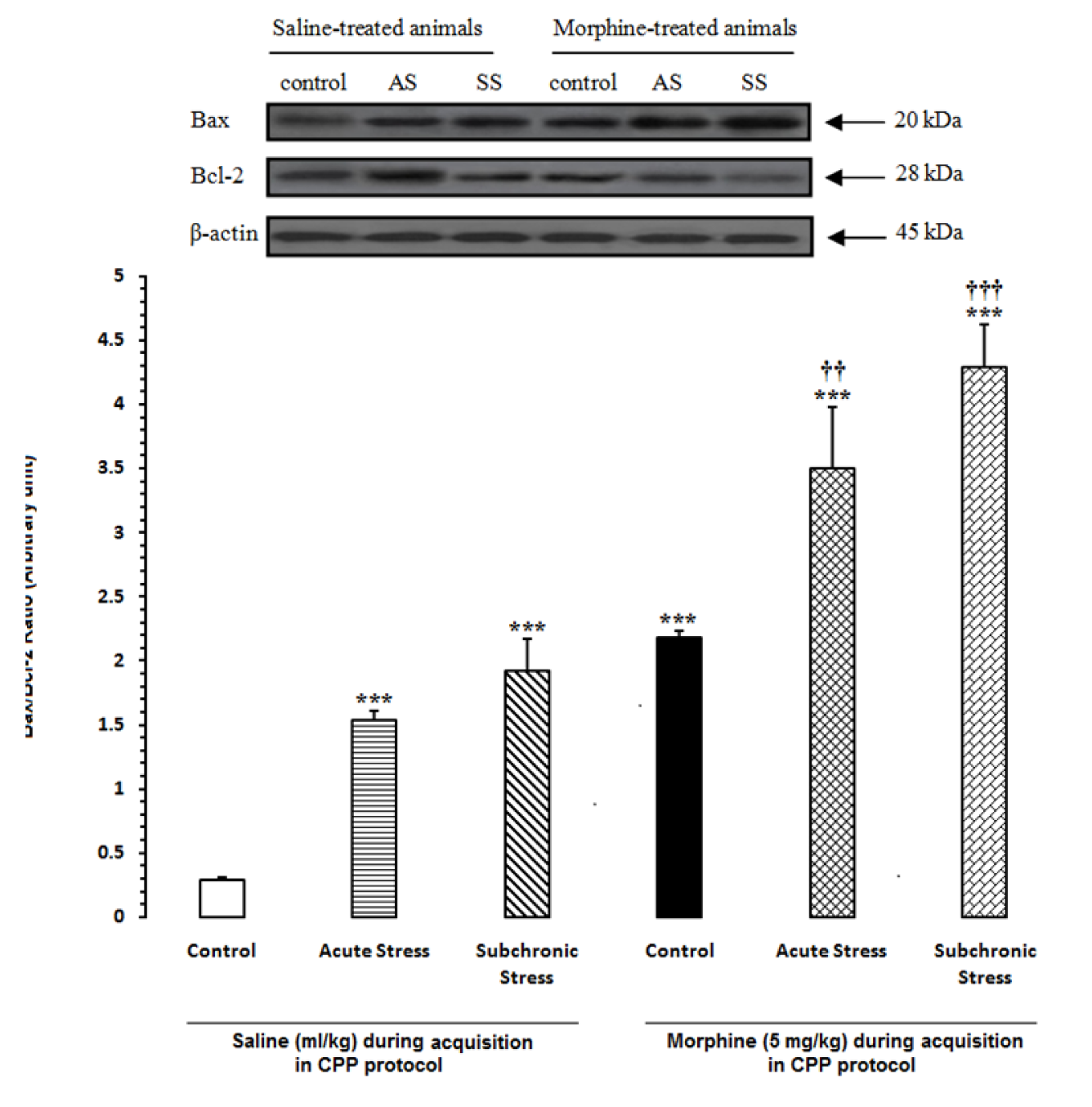
Figure 2: Alteration of Bax/Bcl-2 ratio in the hypothalamus (HYP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean Bax/Bcl-2 ratio calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
*** P<0.001 different from the saline-control group
†† P<0.01, ††† P<0.001 different from the morphine-control group
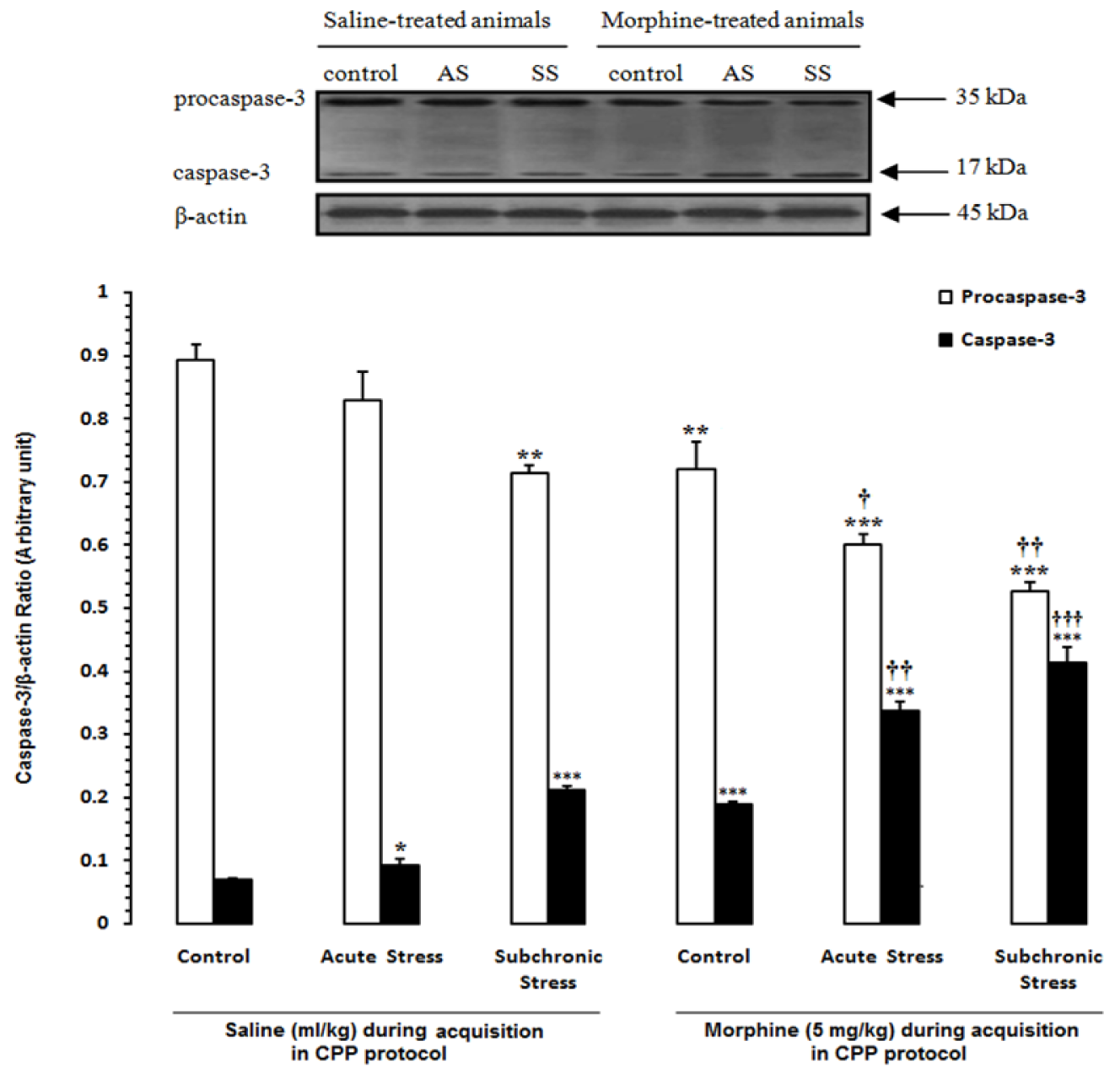
Figure 3: Alteration of caspase-3 in the hypothalamus (HYP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean procaspase-3 and caspase-3 calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
* P<0.05, ** P<0.01,
*** P<0.001 different from the saline-control group
† P<0.05, †† P<0.01,
††† P<0.001 different from the morphine-control group
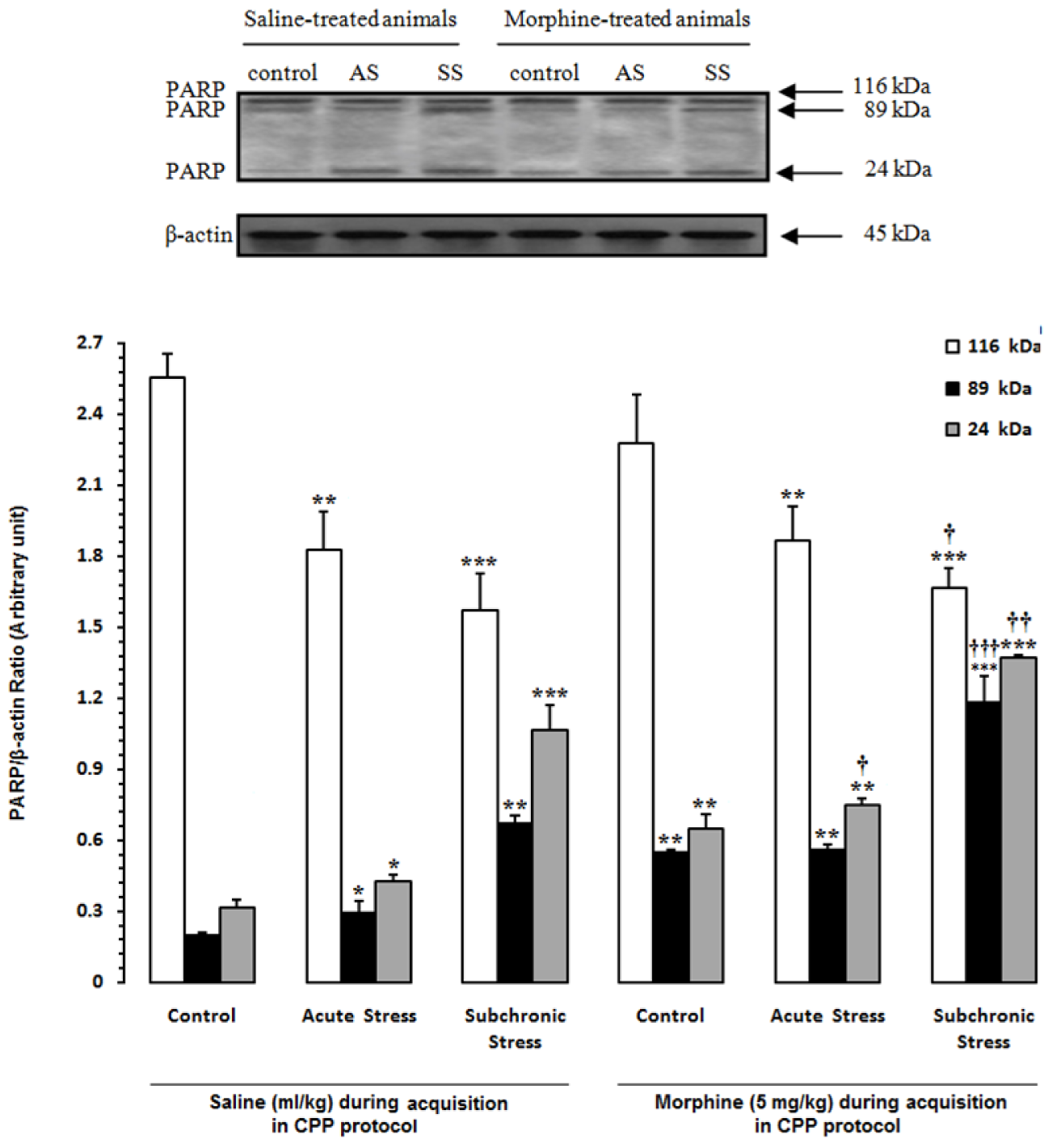
Figure 4: Alteration of PARP in the hypothalamus (HYP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean 116-kDa, 89-kDa and 24-kDa PARPs calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
* P<0.05,** P<0.01,
*** P<0.001 different from the saline-control group
† P<0.05, †† P<0.01, ††† P<0.001 different from the morphine-control group
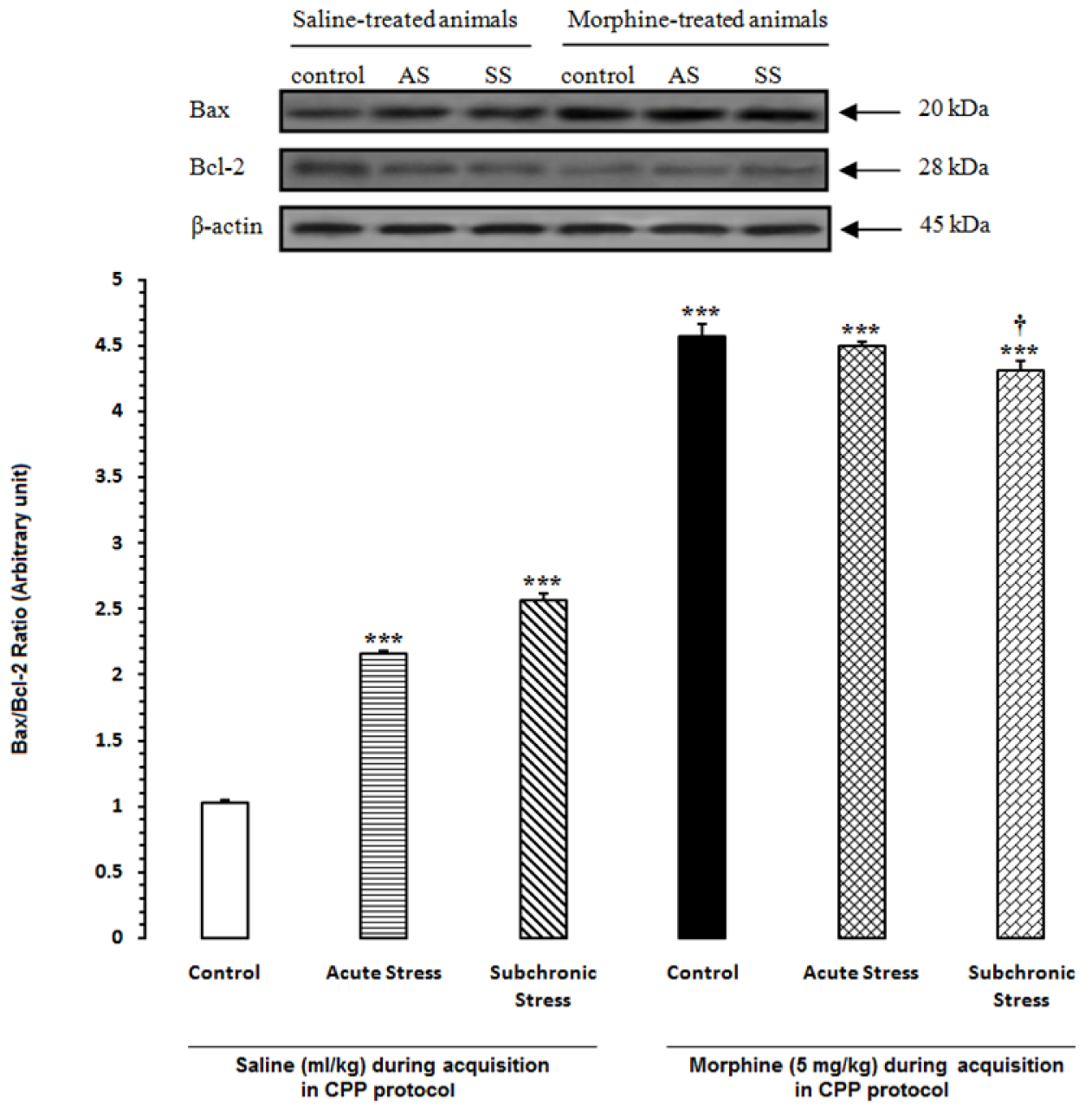
Figure 5: Alteration of Bax/Bcl-2 ratio in the hippocampus (HIP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean Bax/Bcl-2 ratio calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
*** P<0.001 different from the saline-control group
† P<0.05 different from the morphine-control group
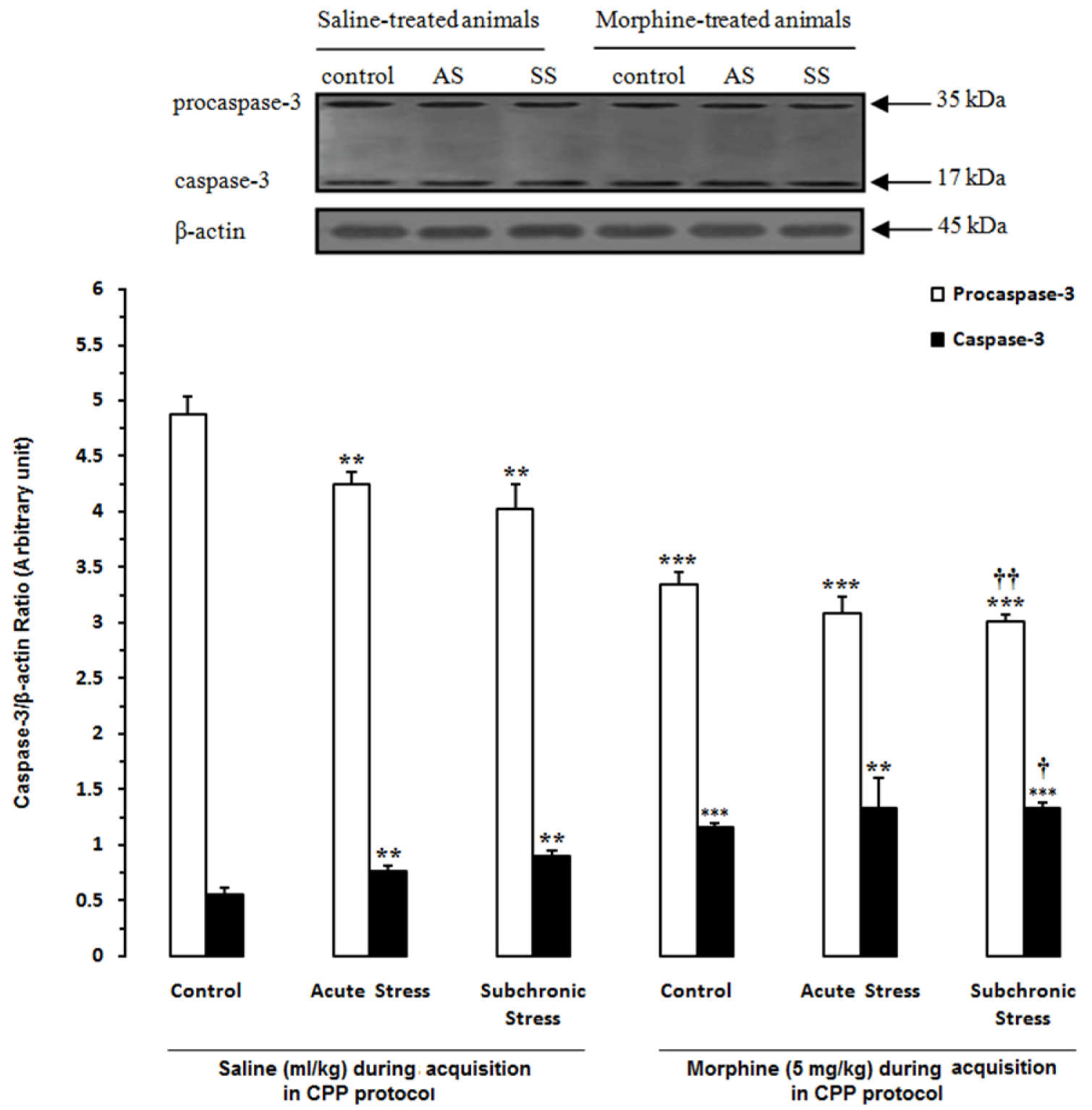
Figure 6: Alteration of caspase-3 in the hippocampus (HIP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean procaspase-3 and caspase-3 calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
** P<0.01,
*** P<0.001 different from the saline-control group
† P<0.05,
†† P<0.01 different from the morphine-control group
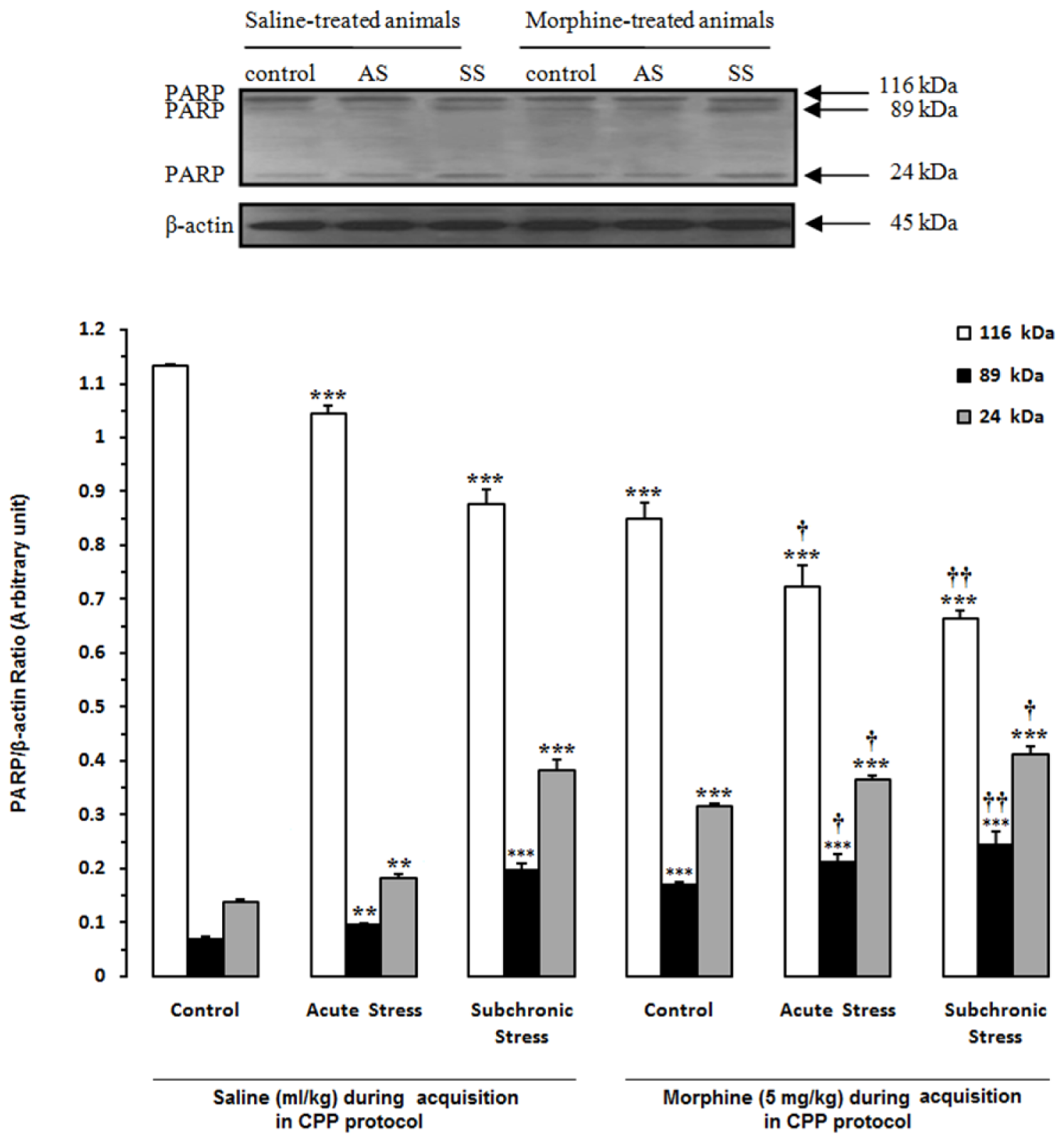
Figure 7: Alteration of PARP in the hippocampus (HIP) after induction of acute and subchronic stress during conditioned place preference paradigm. Upper panels are the representative immunoblots of proteins in this area. Bottom panels show the mean 116-kDa, 89-kDa and 24-kDa PARPs calculated from densitometric quantification of the corresponding bands from left to right, respectively. Each point shows the mean ± SEM for 3-4 rats.
** P<0.01,
*** P<0.001 different from the saline-control group
† P<0.05,
†† P<0.01 different from the morphine-control group
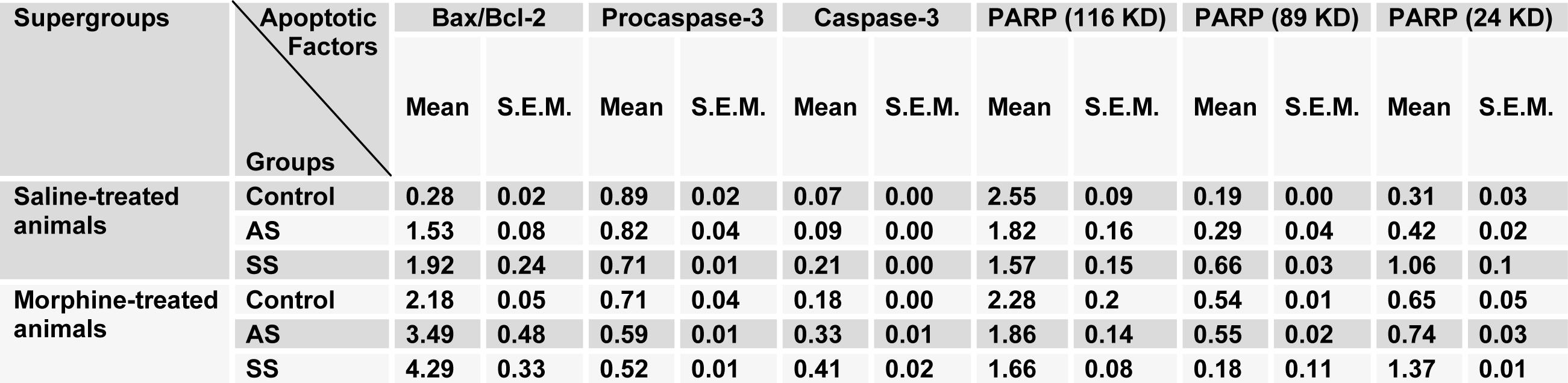
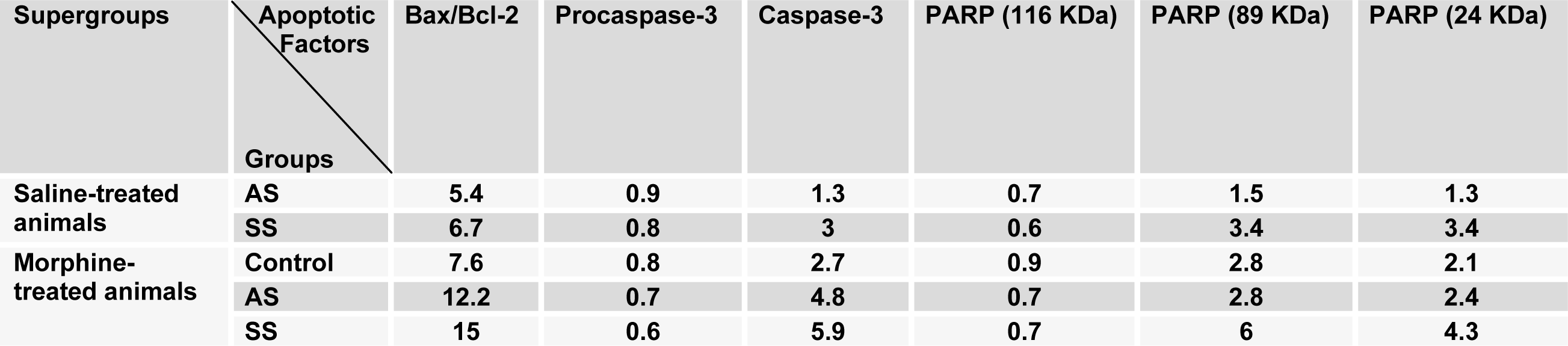
Table 2: The alterations of degree for apoptotic factors in the hypothalamus area compared to control saline-treated animals
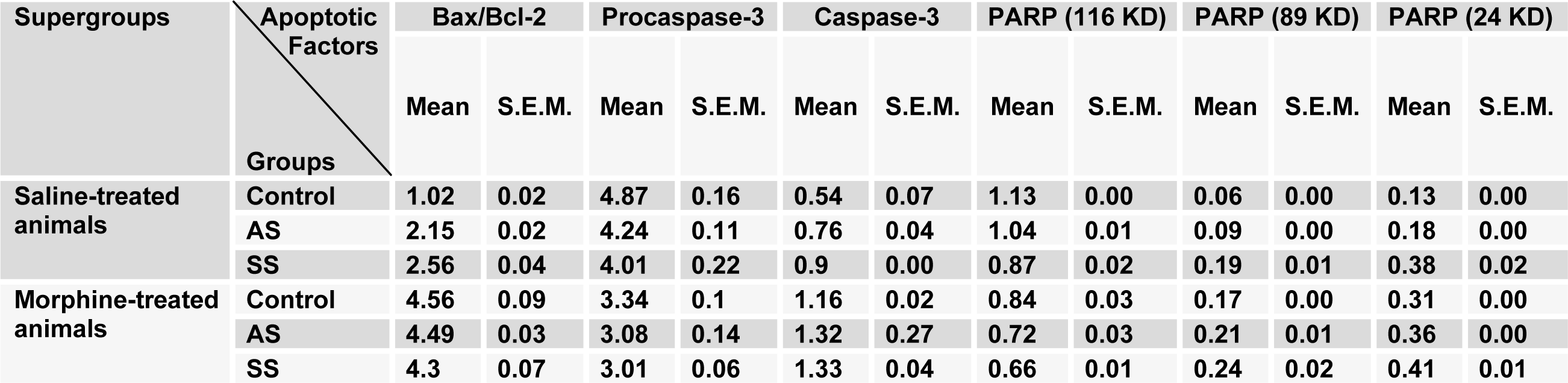
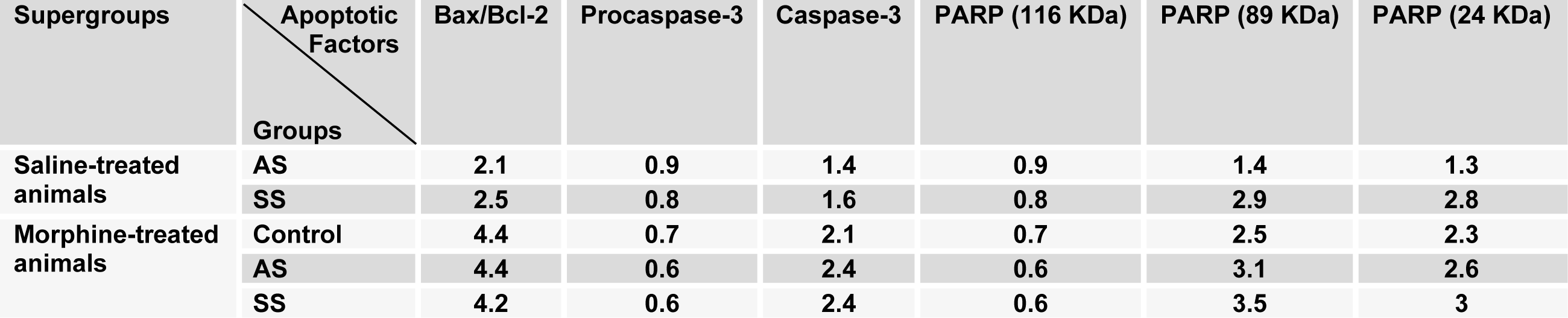
Table 4: The alterations of degree for apoptotic factors in the hippocampus area compared to control saline-treated animals
[*] Corresponding Author:
Ph.D. Abbas Haghparast, Neuroscience Research Center, Shahid Beheshti University of Medical Sciences, P.O. Box 19615-1178, Tehran, Iran; Tel./Fax: +98-21-2243-1624, eMail: Haghparast@yahoo.com